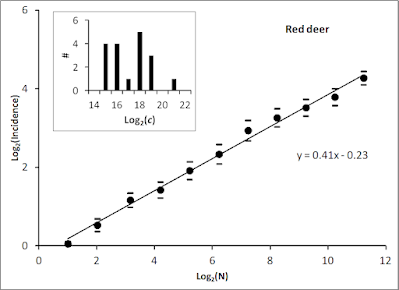
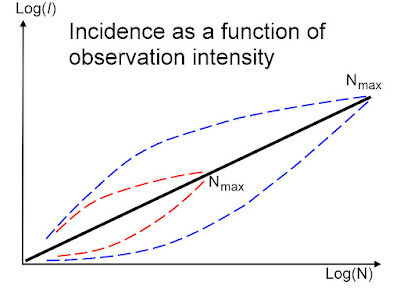
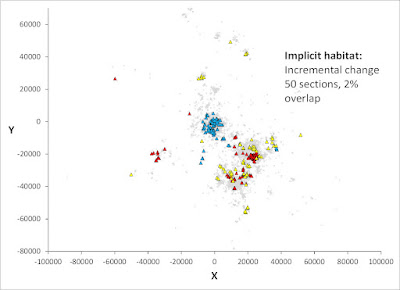
What Rubik's cube teaches us about animal space use

You may cheat the challenge from Rubik's cube by solving it through just seven auto-pilot steps, but taking the painstaking intellectual path without guidance is potentially million of times harder! However, in fact you will then (perhaps unintentionally) stretch your mental capacity and drastically improving your odds by invoking parallel processing. In this post I argue that this "trick" is very similar to the basic principles I'm touting by advocating Multi-scaled Random Walk (MRW), underlying the animal space use model. Deep thinking by kestrel. Photo: AOG. Theoretically, there are 43,252,003,274,489,856,000 (43 quintillion) possible combinations of Rubik moves if you do it by random steps. Mathematically, this approach satisfies a first-order Markovian process. In other words, the future is independent of the past, given the present. As I have repeatedly stated in my book, papers and in this blog, the standard theory of animal space use...