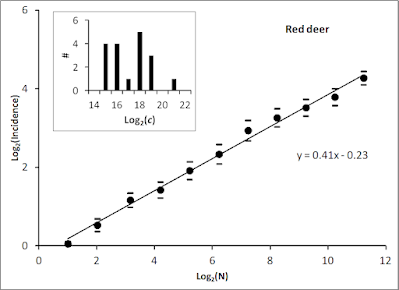
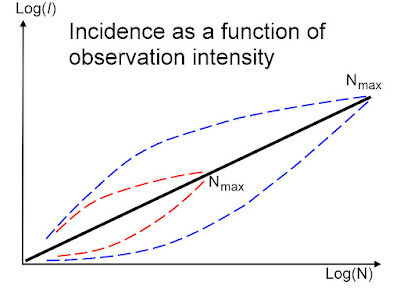
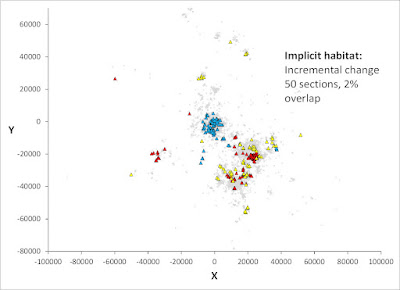
Deeper realism is needed in modelling of population dynamics

Among vertebrates, most populations represent rare species. If this fact is not challenging enough in a survival perspective, all populations - whether rare or abundant - live in an open environment. In some previous posts I've focused on survival-enhancing mechanisms like intraspecific cohesion from conspecific attraction. I have also pointed out the survival value of deterministic individual returns to a homestead following temporary long range excursions outside a given local population. Of course, such lifelines require individuals' cognitive capacity to relate to the environment an explicit spatio-temporal manner. In other words, to counteract the destructive effects of free dispersal beyond the local population's respective core areas, memory map utilization is key. Dispersal is an important aspect of animal space use optimization, but so is the advantage to returning home even over vast distances if this improves survival. If the memory premise becomes broadly accep...