The Biophysical Framework’s Potential for Behavioural Ecology

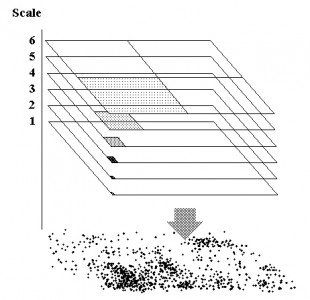
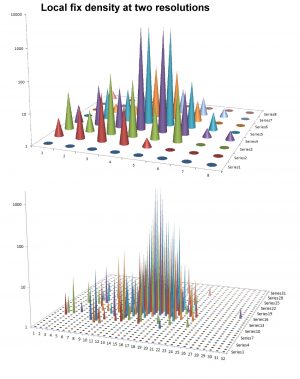
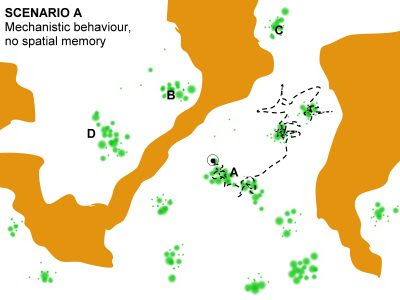
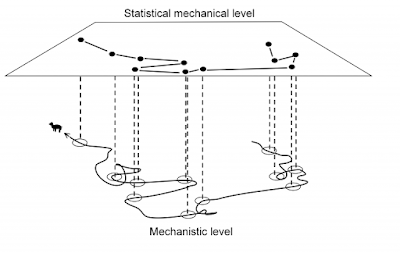
Data collection of individual movement like a series of GPS fixes provides a potential for a physical – a statistical-mechanical – interpretation of animal space use. Such material represents indirect studies of behaviour in contrast to direct observation and interpretation. The GPS pattern of dots on the map provides a coarse-grained image of how the individual in overall terms relocated itself during the period of sampling. It is fascinating that this “out of focus” image may in fact not only be scrutinized with respect to verifying many similar behavioural traits as traditionally studied by ethological methods, but also allows for interpretation of specific relationships that are difficult or outright impossible to test from the classic methods in behavioural ecology. In this post I’m focusing on one of these space use properties, scale-free habitat utilization. First, what is “scale-free movement”? Statistically, this property apparently should be easy to verify (or falsify) by...