MRW and Ecology – Part V: Black Bear Home Ranges Revisited

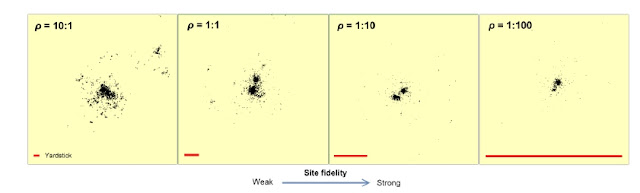
Back in 1994 I enjoyed an unforgettable and extremely inspiring 2-month stay at University of Tennessee, visiting professor Stuart L. Pimm (Department of Ecology and Evolutionary Biology) and Professor Mike L. Pelton (Department of Forestry, Wildlife and Fishery). During some hectic weeks I worked on transforming the mathematical formulation of the Zoomer model for complex population dynamics into a spatially explicit simulation model (Stuart’s lab) in parallel with interaction with many dedicated students of the biology and ecology of black bear Ursus americanu s (Mike’s lab).The Zoomer model is published in my book and already commented on this blog. Regarding the stay at Mike’s lab we published a test on the bears’ general space use, where we found close compliance with the Multi-scaled random walk model, MRW (Gautestad et al. 1998). In this post I revisit the black bear data and find this model’s additional potential to cast light on behavioural ecology in a wildlife management c...