The Binary Home Range

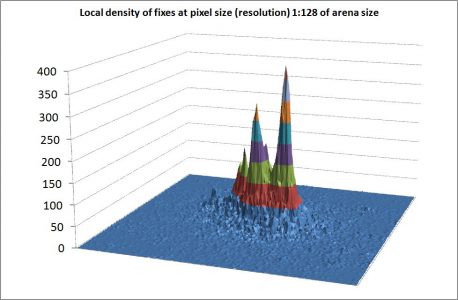
In my foregoing post I criticized the Burt legacy (Burt 1943) for hampering progress in analysis of animal space use on a local scale. Analyses of the spatial pattern of GPS fixes – when this has been explicitly explored by multi-scale methods – consistently confirm a statistical fractal with dimension 0.9 < D < 1.2 rather than a paradigm-confirming to-dimensional area demarcation (albeit with fuzzy borders; see below). In an ideal world the D≈1 result should lead to hefty follow-up tests from the community of animal ecologists for the sake of verifying or falsifying the “home range as a fractal”-model and its behavioural-ecological implications. After all, the home range concept is a cornerstone of animal ecology. Nope. The Burt legacy still appears impenetrable. However, things finally seem to start rolling. After a quarter of a century long invitation period following the initial papers on the topic (Loehle 1990; Gautestad and Mysterud 1993, 1994) the application of fractal...