Positive and Negative Feedback Part II: Populations
Examples of positive feedback loops in population dynamics abound. Even if the majority of models are focusing on negative feedback, like the logistic growth function, non-equilibrium “boom and bust” kind of model designs have also been developed. In this post I elaborate on the particular kind of positive feedback loop that emerges from cross-scale dual-direction flow of individuals that is based on the parallel processing conjecture.
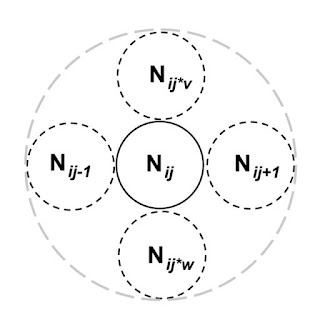 The image to the right illustrates – in simplistic terms – a spatially extended population model of standard kind (e.g., a coupled map lattice design) where each virtually demarcated local population j at spatial resolution i and at a given point in time t contains Nij individuals. No borders for local migration are assumed; i.e., the environment is open both internally and externally towards neighbour sites.Typically, these individuals are set to be subject to a locally negative feedback loop in accordance to principles of density dependent regulation*. The larger the N the larger the probability of an increased death rate and/or and increased emigration rate from time t to t+1, eventually leading both the local and the over-all population to a steady state. This balancing** condition lasts until some change (external perturbation) is forcing the system into a renewed loop of negative feedback-driven dynamics. In a variant of this design, density regulation may be formulated to be absent until a critical local density is reached, leading to boom and bust (“catastrophic” death and emigration), which may be more or less perturbed by random immigration rate from asynchronous developments in respective surrounding Nij. More sophisticated variants abound, like inclusion of time lag responses, interactions with other trophic levels, and so on.
The image to the right illustrates – in simplistic terms – a spatially extended population model of standard kind (e.g., a coupled map lattice design) where each virtually demarcated local population j at spatial resolution i and at a given point in time t contains Nij individuals. No borders for local migration are assumed; i.e., the environment is open both internally and externally towards neighbour sites.Typically, these individuals are set to be subject to a locally negative feedback loop in accordance to principles of density dependent regulation*. The larger the N the larger the probability of an increased death rate and/or and increased emigration rate from time t to t+1, eventually leading both the local and the over-all population to a steady state. This balancing** condition lasts until some change (external perturbation) is forcing the system into a renewed loop of negative feedback-driven dynamics. In a variant of this design, density regulation may be formulated to be absent until a critical local density is reached, leading to boom and bust (“catastrophic” death and emigration), which may be more or less perturbed by random immigration rate from asynchronous developments in respective surrounding Nij. More sophisticated variants abound, like inclusion of time lag responses, interactions with other trophic levels, and so on.
As previously explained in other posts, this kind of model framework depends on a premise of Markov-compliant processes at the individual level (mechanistic system), and thus also at the population level (local or global compliance with the mean field principle). In this framework intrinsic dynamics may be density dependent or not, but from the perspective of a given Nij, extrinsic influence – like immigration of individuals – is always stochastic and thus density independent with respect to Nij. In other words, the net immigration rate during a given time increment is not influenced by the state of the population in this location (i,j). You can search my blog or read my book to find descriptions and details on all these concepts.
To implement cross-location and dual-direction deterministic dynamics, multi-scaled behaviour and spatial memory needs to be introduced. My parallel processing conjecture; which spins off various testable hypotheses, creates turmoil in this standard system design for population dynamics because it explicitly introduces such system complexity. For example, positive feedback loops may emerge. Positive feedback as described below may effectively also counteracting the paradoxical Allée effect, which all “standard” population models are confronted with at the border zone of a population in an open environment**.
The dynamic driver of the complexity is the introduction of spatial memory in combination with a scale-free kind of dynamics along both the spatial and the temporal dimensions. In statistical-mechanical terms, parallel processing is incompatible with a mechanistic system. Thus, a kind of extended statistical mechanics is needed. I refer to the post where I describe the scale-extended description of a metapopulation system.
For the most extensive individual-level test of the parallel processing conjecture until now (indirectly also verifying positive feedback of space use), see our paper on statistical analysis of space use by red deer Cervus elaphus (Gautestad et al. 2013; Gautestad and Mysterud 2013). In my blog I have also provided several anecdotal examples of third party research potentially supporting the parallel processing conjecture. For the sake of system coherence, if parallel processing is verified for individual space use of a given species and under given ecological conditions, this behaviour should also be reflected in the complementary population dynamical modelling of the given species and conditions.
Extending the standard population model. As explained in a range of blog posts, my Zoomer model represents a population level system design that is coherent with the individual-level space use process (in parsimonious terms), as formulated by the Multi-scaled random walk model. In my previous post I described the latter in the context of positive feedback from individual-level site fidelity. Below I illustrate positive feedback also at the population level, where site fidelity get boosted by conspecific attraction. In other words, conspecifics become part of the individuals’ resource mapping at coarser scales, as it is allowed for by spatial memory. Consequently, a potential for dual-direction deterministic flow of individuals is introduced (see above). Conspecific attraction is assumed to be gradually developed by individual experience of conspecifics’ whereabouts during exploratory moves.
First, consider the zooming process, whereby a given rate, z, of individuals (for example, z=5% on average at a chosen time resolution Δt) at a “unit” reference scale (k=i) are redistributing themselves over a scale range beyond this unit scale***. During a given Δt consider that 100 individuals become zoomers from the specific location marked by the white circle. In parallel with the zooming out-process the model describes a zooming in-process with a similar strength. The latter redistributes the zoomers in accordance to scale-free immigration of individuals under conspecific attraction.Thus, number of individuals (N) at this location j at scale i, marked as Nij, will at the next time t+1 either embed N-100 individuals if they all leave location j and end up somewhere in the neighbourhood of j, or the new number will be N -100 + an influx of immigrants, where these immigrants come from the neighbourhood at scale i (those returning home again), scale i+1 (immigration from locations nearby), i+2 (from an even more distant neighbourhood), etc.

Image above: In the Zoomer model , some percentage of the individuals are redistributing themselves over a scale range during each time increment. Emigration (“zooming out”) is marked by dotted arrows, and immigration (“zooming in”) is shown as continuous-line arrows. Numbers refer to scale level of the neighbourhood of a given locality. This neighbourhood scales logarithmically; i.e., in a scale-free manner, in compliance with exploratory moves in the individual-level Multi-scaled random walk model. Zooming in depends on spatial memory by the individuals, and introduces a potential for the emergence of positive feedback at the population level.
In the ideal model variant of zooming we are thus assuming a scale-free redistribution of individuals during zooming, with zooming to a neighbourhood at scale ki+x takes place with probability 1/ki+x (Gautestad and Mysterud 2005). Under this condition, zoomers to successively coarser scales become “diluted” over proportionally larger neighbourhood area, the maximum number of immigrants in this example is 100 + N’, where N’ is the average number of zoomers pr. location at unit scale k=i within the coarsest defined system scale k=i(max) for zooming surrounding location j at scale i.
As a consequence of this kind of scale-free emigration of zoomers, the population system demonstrates zooming with equal weight of individual redistribution from scale to scale over the defined scale range (Lévy-like in this respect, with scaling exponent β≈2; see Gautestad and Mysterud 2005). By studying the distribution of step lengths, this “equal weight” hypothesis may be tested, when combinded with other statistical fingerprints (in particular, verifying memory-dependent site fidelity; see Gautestad and Mysterud 2013).
Putting this parsimonious Zoomer model with its system variables and parameters into a specific ecological context implies a huge and basically unexplored potential for ecological inference under condition of scale-free space use in combination with site fidelity.
Positive feedback in the Zoomer model. As shown in my series of simulations of the Zoomer model a few posts ago, a positive feedback loop emerges from locations with relatively high abundance of individuals having a relatively larger chance of received a net influx of zoomers during the next increment, and vice versa for locations with low abundance. The positive feedback emerges from the conspecific attraction process, linking the dynamics at different scales together in a parallel processing manner.
This positive feedback loop from conspecific attraction also counteracts extinction from a potential Allée effect (see this post and this post), which have traditionally been understood and formulated from the standard population paradigm. The Zoomer model represents an alternative description of a process that effectively counteracts this effect.
NOTES
*) The migration rates connects the local population to surrounding populations. Immigration is – by necessity from the standard model design – density independent with respect to the dynamics in Nij.
**) Since the process is assumed to obey a Markovian and the mean field principles (standard, mechanistic process), the arena and population system must either be assumed to be infinitely large or the total set of local populations has to be assumed to be demarcated by some kind of physical border. Otherwise, net emigration and increased death rate in the border zone will tend to drive N towards zero in open environments (extinction from standard diffusion in combination with local N drifting below critical density where Allée kicks in). Individuals will “leak” from an open border zone to the surroundings where N is lower.
***) The unit temporal scale for a population system should be considered coarser than the unit scale at the individual level, since the actual scale range under scrutiny typically is larger for population systems. In particular, to find the temporal scale where for example 5% of the local population can be expected to be moving past the inter-cell borders of a given unit spatial grid resolution ki=1, one should be expected to find Δt substantially larger than Δt at the individual level.
Consider that the difference in Δt is a function of the difference of the area of short-range versus long range displacements under the step length curve for individual displacements, where the ∼5% long-step tail of this area represents the relative unit time in comparison to the rest of the distribution (thereby defined as intra-cell moves). Since this area is a fraction of the area for the remaining 95% of the displacements, the difference in Δt should scale accordingly.
REFERENCES
Gautestad, A. O., and I. Mysterud. 2005. Intrinsic scaling complexity in animal dispersion and abundance. The American Naturalist 165:44-55.
Gautestad, A. O., and A. Mysterud. 2013. The Lévy flight foraging hypothesis: forgetting about memory may lead to false verification of Brownian motion. Movement Ecology 1:1-18.
Gautestad, A. O., L. E. Loe, and A. Mysterud. 2013. Inferring spatial memory and spatiotemporal scaling from GPS data: comparing red deer Cervus elaphus movements with simulation models. Journal of Animal Ecology 82:572-586.

As previously explained in other posts, this kind of model framework depends on a premise of Markov-compliant processes at the individual level (mechanistic system), and thus also at the population level (local or global compliance with the mean field principle). In this framework intrinsic dynamics may be density dependent or not, but from the perspective of a given Nij, extrinsic influence – like immigration of individuals – is always stochastic and thus density independent with respect to Nij. In other words, the net immigration rate during a given time increment is not influenced by the state of the population in this location (i,j). You can search my blog or read my book to find descriptions and details on all these concepts.
To implement cross-location and dual-direction deterministic dynamics, multi-scaled behaviour and spatial memory needs to be introduced. My parallel processing conjecture; which spins off various testable hypotheses, creates turmoil in this standard system design for population dynamics because it explicitly introduces such system complexity. For example, positive feedback loops may emerge. Positive feedback as described below may effectively also counteracting the paradoxical Allée effect, which all “standard” population models are confronted with at the border zone of a population in an open environment**.
The dynamic driver of the complexity is the introduction of spatial memory in combination with a scale-free kind of dynamics along both the spatial and the temporal dimensions. In statistical-mechanical terms, parallel processing is incompatible with a mechanistic system. Thus, a kind of extended statistical mechanics is needed. I refer to the post where I describe the scale-extended description of a metapopulation system.
For the most extensive individual-level test of the parallel processing conjecture until now (indirectly also verifying positive feedback of space use), see our paper on statistical analysis of space use by red deer Cervus elaphus (Gautestad et al. 2013; Gautestad and Mysterud 2013). In my blog I have also provided several anecdotal examples of third party research potentially supporting the parallel processing conjecture. For the sake of system coherence, if parallel processing is verified for individual space use of a given species and under given ecological conditions, this behaviour should also be reflected in the complementary population dynamical modelling of the given species and conditions.
Extending the standard population model. As explained in a range of blog posts, my Zoomer model represents a population level system design that is coherent with the individual-level space use process (in parsimonious terms), as formulated by the Multi-scaled random walk model. In my previous post I described the latter in the context of positive feedback from individual-level site fidelity. Below I illustrate positive feedback also at the population level, where site fidelity get boosted by conspecific attraction. In other words, conspecifics become part of the individuals’ resource mapping at coarser scales, as it is allowed for by spatial memory. Consequently, a potential for dual-direction deterministic flow of individuals is introduced (see above). Conspecific attraction is assumed to be gradually developed by individual experience of conspecifics’ whereabouts during exploratory moves.
First, consider the zooming process, whereby a given rate, z, of individuals (for example, z=5% on average at a chosen time resolution Δt) at a “unit” reference scale (k=i) are redistributing themselves over a scale range beyond this unit scale***. During a given Δt consider that 100 individuals become zoomers from the specific location marked by the white circle. In parallel with the zooming out-process the model describes a zooming in-process with a similar strength. The latter redistributes the zoomers in accordance to scale-free immigration of individuals under conspecific attraction.Thus, number of individuals (N) at this location j at scale i, marked as Nij, will at the next time t+1 either embed N-100 individuals if they all leave location j and end up somewhere in the neighbourhood of j, or the new number will be N -100 + an influx of immigrants, where these immigrants come from the neighbourhood at scale i (those returning home again), scale i+1 (immigration from locations nearby), i+2 (from an even more distant neighbourhood), etc.

Image above: In the Zoomer model , some percentage of the individuals are redistributing themselves over a scale range during each time increment. Emigration (“zooming out”) is marked by dotted arrows, and immigration (“zooming in”) is shown as continuous-line arrows. Numbers refer to scale level of the neighbourhood of a given locality. This neighbourhood scales logarithmically; i.e., in a scale-free manner, in compliance with exploratory moves in the individual-level Multi-scaled random walk model. Zooming in depends on spatial memory by the individuals, and introduces a potential for the emergence of positive feedback at the population level.
In the ideal model variant of zooming we are thus assuming a scale-free redistribution of individuals during zooming, with zooming to a neighbourhood at scale ki+x takes place with probability 1/ki+x (Gautestad and Mysterud 2005). Under this condition, zoomers to successively coarser scales become “diluted” over proportionally larger neighbourhood area, the maximum number of immigrants in this example is 100 + N’, where N’ is the average number of zoomers pr. location at unit scale k=i within the coarsest defined system scale k=i(max) for zooming surrounding location j at scale i.
As a consequence of this kind of scale-free emigration of zoomers, the population system demonstrates zooming with equal weight of individual redistribution from scale to scale over the defined scale range (Lévy-like in this respect, with scaling exponent β≈2; see Gautestad and Mysterud 2005). By studying the distribution of step lengths, this “equal weight” hypothesis may be tested, when combinded with other statistical fingerprints (in particular, verifying memory-dependent site fidelity; see Gautestad and Mysterud 2013).
Putting this parsimonious Zoomer model with its system variables and parameters into a specific ecological context implies a huge and basically unexplored potential for ecological inference under condition of scale-free space use in combination with site fidelity.
Positive feedback in the Zoomer model. As shown in my series of simulations of the Zoomer model a few posts ago, a positive feedback loop emerges from locations with relatively high abundance of individuals having a relatively larger chance of received a net influx of zoomers during the next increment, and vice versa for locations with low abundance. The positive feedback emerges from the conspecific attraction process, linking the dynamics at different scales together in a parallel processing manner.
This positive feedback loop from conspecific attraction also counteracts extinction from a potential Allée effect (see this post and this post), which have traditionally been understood and formulated from the standard population paradigm. The Zoomer model represents an alternative description of a process that effectively counteracts this effect.
NOTES
*) The migration rates connects the local population to surrounding populations. Immigration is – by necessity from the standard model design – density independent with respect to the dynamics in Nij.
**) Since the process is assumed to obey a Markovian and the mean field principles (standard, mechanistic process), the arena and population system must either be assumed to be infinitely large or the total set of local populations has to be assumed to be demarcated by some kind of physical border. Otherwise, net emigration and increased death rate in the border zone will tend to drive N towards zero in open environments (extinction from standard diffusion in combination with local N drifting below critical density where Allée kicks in). Individuals will “leak” from an open border zone to the surroundings where N is lower.
***) The unit temporal scale for a population system should be considered coarser than the unit scale at the individual level, since the actual scale range under scrutiny typically is larger for population systems. In particular, to find the temporal scale where for example 5% of the local population can be expected to be moving past the inter-cell borders of a given unit spatial grid resolution ki=1, one should be expected to find Δt substantially larger than Δt at the individual level.
Consider that the difference in Δt is a function of the difference of the area of short-range versus long range displacements under the step length curve for individual displacements, where the ∼5% long-step tail of this area represents the relative unit time in comparison to the rest of the distribution (thereby defined as intra-cell moves). Since this area is a fraction of the area for the remaining 95% of the displacements, the difference in Δt should scale accordingly.
REFERENCES
Gautestad, A. O., and I. Mysterud. 2005. Intrinsic scaling complexity in animal dispersion and abundance. The American Naturalist 165:44-55.
Gautestad, A. O., and A. Mysterud. 2013. The Lévy flight foraging hypothesis: forgetting about memory may lead to false verification of Brownian motion. Movement Ecology 1:1-18.
Gautestad, A. O., L. E. Loe, and A. Mysterud. 2013. Inferring spatial memory and spatiotemporal scaling from GPS data: comparing red deer Cervus elaphus movements with simulation models. Journal of Animal Ecology 82:572-586.