Fractal Compliant Space Use: Intrinsic Scaling or Extrinsic Habitat Heterogeneity?

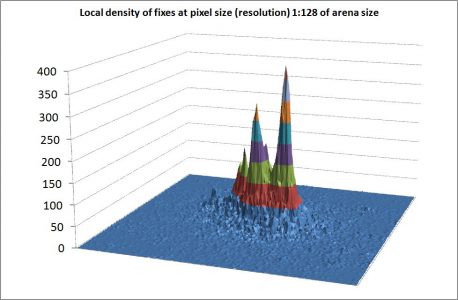
A growing number of analyses of spatial scatter of GPS positions of animals (fixes) verify a fractal dimension (D) substantially smaller than what should be expected from standard models. Specifically, D tends to be close to 1, which reflects a very heterogeneous fix dispersion over a wide range of spatial resolutions. A recurring question among analysts of animal space use is thus: is this so-called scale-free pattern – statistically speaking – reflecting a matching heterogeneous habitat that happens to satisfy a self-similar dispersion (the “environmental forcing” explanation), or is the scale-free space use a manifestation of intrinsic, cognitive processes (the “emergent property” explanation)? In this post I illustrate the challenge by showing an analysis of large mammals' space use. The image above, showing the spatial accumulation of fixes from GPS-sampling a red deer Cervus elaphus individual during the summer season illustrates this key question (see Gautestad et al. 2...